

Abstract
Pediatric AML has a relapse rate approaching 40%, indicating that resistance to chemotherapy remains a critical problem. We are focused on identifying and overcoming environment-mediated mechanisms of resistance. We previously reported that the sensitivity of G-CSF- and IL-6-induced STAT3 signaling was significantly associated with outcome in pediatric AML patients. In this follow-up study, we used conditioned medium (CM) from HS5 stromal cells as a more physiological stimulus to evaluate the capacity of primary AML cells to activate signaling pathways.
We studied 111 diagnostic bone marrow samples from patients who enrolled on Children's Oncology Group (COG) AML treatment studies, AAML03P1 or AAML0531, and provided informed consent to bank bone marrow. The same chemotherapy backbone was used in both trials. All samples had at least 70% viability after thawing. Thawed cells were divided into aliquots for stimulation with 50% CM, 10 ng/ml G-CSF, or 5 ng/ml IL-6. Unstimulated cells were used for isotype controls and for determination of basal signaling levels. Following 15 min stimulation, cells were fixed and processed for FACS analysis of CD45, pY-STAT3, pY-STAT5, pERK1/2, and pAKT. Responses were expressed as the fold change in mean fluorescence intensity for stimulated cells over unstimulated cells (ΔMFI).
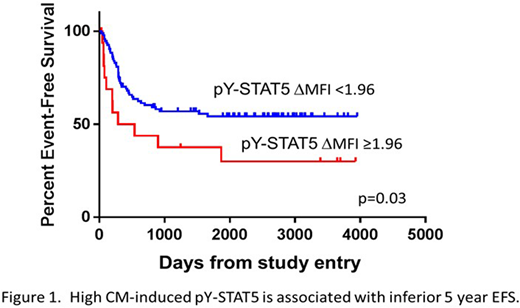
The ΔMFIs for pY-STAT3 and pY-STAT5 varied between 0.1 and 31.8. The ΔMFIs for pERK1/2 and pAKT rarely exceeded 2. Samples with a robust response to one stimulus generally responded robustly to the others. The Pearson correlation coefficient (r) for CM-induced pY-STAT3 v. G-CSF-induced pY-STAT3 was 0.8511 (p<0.00001). The CM-induced pY-STAT5 and G-CSF-induced pY-STAT5 responses were significantly but less strongly correlated (r=0.2765; p=0.0043). Cut point analyses identified response thresholds that distinguished patients with higher EFS from those with lower EFS. We found that higher CM-induced pY-STAT5 was significantly associated with an inferior EFS (Figure 1), with HR 2.06 for those with ΔMFI > 1.96 (p=0.034). Previously we found that higher inducible pY-STAT3 was associated with significantly better EFS (Redell, et al, Blood, 2013; Long, et al, Oncotarget, 2017). In this study, no cut point for CM-induced pY-STAT3 ΔMFI distinguished patients with good or poor EFS. Cytogenetics and FLT3/ITD were not significantly different for groups above and below the ΔMFI cut points. The finding that robust STAT5 signaling is associated with inferior EFS suggests that a factor that signals primarily through STAT5 and not STAT3 (e.g. GM-CSF, IL-3, FL) could contribute to treatment resistance and relapse.
Interestingly, most of the samples that failed to activate STAT3/5 pathways in response to G-CSF also failed to respond to the cocktail of factors in CM, suggesting a generalized signaling dysfunction. To investigate intrinsic gene expression differences, we leveraged existing RNA-seq data for the samples for which we generated signaling responses. We obtained RNA-seq data for 27 samples from the NCI Therapeutically Applicable Research to Generate Effective Treatments (TARGET) database. RNA-seq data for an additional 45 samples were provided through collaboration with the University of Washington. Of these 72 samples, 29 samples were resistant to CM-induced pY-STAT3 (ΔMFI <2, CM-R) and 43 samples were sensitive (ΔMFI > 2, CM-S). Raw counts were normalized and analyzed by EdgeR with GLM algorithm and blocking to control for batch effects from the two datasets. We found that 219 features were significantly differentially expressed (DE; FDR <0.05), with 192 being expressed more highly in the CM-R group. Gene set enrichment analysis identified the "matrisome" gene set, including genes encoding growth factors and matrix proteins, as being significantly enriched in the CM-R group. For example, genes encoding G-CSF, GM-CSF, WNT7B, and integrin B3 were upregulated in CM-R samples. Additionally, a number of non-HOX homeobox genes, including DLX2, DLX3 and MSX2, were increased in CM-R samples. There was no difference in the mean %blasts for the samples in the CM-R and CM-S groups, arguing against the increased expression of matrix-related genes being due to a higher proportion of non-blast cells in the CM-R samples. Our integration of gene expression with inducible STAT3/5 responses will yield novel insights into extrinsic survival signaling and mechanisms of dysfunction.
No relevant conflicts of interest to declare.

This feature is available to Subscribers Only
Sign In or Create an Account Close Modal